Биосинтез белков в клетках представляет собой последовательность реакций матричного типа, в ходе которых последовательная передача наследственной информации с одного типа молекул на другой приводит к образованию полипептидов с генетически обусловленной структурой.
Биосинтез белков представляет собой начальный этап реализации, или экспрессии генетической информации. К главным матричным процессам, обеспечивающим биосинтез белков, относятся транскрипция ДНК и трансляция мРНК. Транскрипция ДНК заключается в переписывании информации с ДНК на мРНК (матричную, или информационную РНК). Трансляция мРНК заключается в переносе информации с мРНК на полипептид. Последовательность матричных реакций при биосинтезе белков можно представить в виде схемы.
нетранскрибируемая цепь ДНК
транскрибируемая цепь ДНК
транскрипция ДНК
кодоны мРНК
трансляция мРНК
антикодоны тРНК
аминокислоты белка
метионин
На схеме видно, что генетическая информация о структуре белка хранится в виде последовательности триплетов ДНК. При этом лишь одна из цепей ДНК служит матрицей для транскрипции (такая цепь называется транскрибируемой). Вторая цепь является комплементарной по отношению к транскрибируемой и не участвует в синтезе мРНК.
Молекула мРНК служит матрицей для синтеза полипептида на рибосомах. Триплеты мРНК, кодирующие определенную аминокислоту, называются кодоны. В трансляции принимают участие молекулы тРНК. Каждая молекула тРНК содержит антикодон – распознающий триплет, в котором последовательность нуклеотидов комплементарна по отношению к определенному кодону мРНК. Каждая молекула тРНК способна переносить строго определенную аминокислоту. Соединение тРНК с аминокислотой называется аминоацил–тРНК.
Молекула тРНК по общей конформации напоминает клеверный лист на черешке. «Вершина листа» несет антикодон. Существует 61 тип тРНК с разными антикодонами. К «черешку листа» присоединяется аминокислота (существует 20 аминокислот, участвующих в синтезе полипептида на рибосомах). Каждой молекуле тРНК с определенным антикодоном соответствует строго определенная аминокислота. В то же время, определенной аминокислоте обычно соответствует несколько типов тРНК с разными антикодонами. Аминокислота ковалентно присоединяется к тРНК с помощью ферментов – аминоацил-тРНК-синтетаз. Эта реакция называется аминоацилированием тРНК.
На рибосомах к определенному кодону мРНК с помощью специфического белка присоединяется антикодон соответствующей молекулы аминоацил-тРНК. Такое связывание мРНК и аминоацил-тРНК называется кодонзависимым. На рибосомах аминокислоты соединяются между собой с помощью пептидных связей, а освободившиеся молекулы тРНК уходят на поиски свободных аминокислот.
Рассмотрим подробнее основные этапы биосинтеза белков.
1 этап. Транскрипция ДНК. На транскрибируемой цепи ДНК с помощью ДНК-зависимой РНК-полимеразы достраивается комплементарная цепь мРНК. Молекула мРНК является точной копией нетранскрибируемой цепи ДНК с той разницей, что вместо дезоксирибонуклеотидов в ее состав входят рибонуклеотиды, в состав которых вместо тимина входит урацил.
2 этап. Процессинг (созревание) мРНК. Синтезированная молекула мРНК (первичный транскрипт) подвергается дополнительным превращениям. В большинстве случаев исходная молекула мРНК разрезается на отдельные фрагменты. Одни фрагменты – интроны – расщепляются до нуклеотидов, а другие – экзоны – сшиваются в зрелую мРНК. Процесс соединения экзонов «без узелков» называетсясплайсинг.
Сплайсинг характерен для эукариот и архебактерий, но иногда встречается и у прокариот. Существует несколько видов сплайсинга. Сущность альтернативного сплайсинга заключается в том, что одни и те же участки исходной мРНК могут быть и интронами, и экзонами. Тогда одному и тому же участку ДНК соответствует несколько типов зрелой мРНК и, соответственно, несколько разных форм одного и того же белка. Сущность транс–сплайсинга заключается в соединение экзонов, кодируемых разными генами (иногда даже из разных хромосом), в одну зрелую молекулу мРНК.
3 этап. Трансляция мРНК. Трансляция (как и все матричные процессы) включает три стадии: инициацию (начало), элонгацию (продолжение) и терминацию (окончание).
Инициация. Сущность инициации заключается в образовании пептидной связи между двумя первыми аминокислотами полипептида.
Первоначально образуется инициирующий комплекс, в состав которого входят: малая субъединица рибосомы, специфические белки (факторы инициации) и специальная инициаторная метиониновая тРНК с аминокислотой метионином – Мет–тРНКМет. Инициирующий комплекс узнает начало мРНК, присоединяется к ней и скользит до точки инициации (начала) биосинтеза белка: в большинстве случаев это стартовый кодон АУГ. Между стартовым кодоном мРНК и антикодоном метиониновой тРНК происходит кодонзависимое связывание с образованием водородных связей. Затем происходит присоединение большой субъединицы рибосомы.
При объединении субъединиц образуется целостная рибосома, которая несет два активных центра (сайта): А–участок (аминоацильный, который служит для присоединения аминоацил-тРНК) и Р–участок (пептидилтрансферазный, который служит для образования пептидной связи между аминокислотами).
Первоначально Мет–тРНКМет находится на А–участке, но затем перемещается на Р–участок. На освободившийся А–участок поступает аминоацил-тРНК с антикодоном, который комплементарен кодону мРНК, следующему за кодоном АУГ. В нашем примере это Гли–тРНКГли с антикодоном ЦЦГ, который комплементарен кодону ГГЦ. В результате кодонзависимого связывания между кодоном мРНК и антикодоном аминоацил-тРНК образуются водородные связи. Таким образом, на рибосоме рядом оказываются две аминокислоты, между которыми образуется пептидная связь. Ковалентная связь между первой аминокислотой (метионином) и её тРНК разрывается.
После образования пептидной связи между двумя первыми аминокислотами рибосома сдвигается на один триплет. В результате происходит транслокация (перемещение) инициаторной метиониновой тРНКМет за пределы рибосомы. Водородная связь между стартовым кодоном и антикодоном инициаторной тРНК разрывается. В результате свободная тРНКМет отщепляется и уходит на поиск своей аминокислоты.
Вторая тРНК вместе с аминокислотой (в нашем примере Гли–тРНКГли) в результате транслокации оказывается на Р–участке, а А–участок освобождается.
Элонгация. Сущность элонгации заключается в присоединении последующих аминокислот, то есть в наращивании полипептидной цепи. Рабочий цикл рибосомы в процессе элонгации состоит из трех шагов: кодонзависимого связывания мРНК и аминоацил-тРНК на А–участке, образования пептидной связи между аминокислотой и растущей полипептидной цепью и транслокации с освобождением А–участка.
На освободившийся А–участок поступает аминоацил-тРНК с антикодоном, соответствующим следующему кодону мРНК (в нашем примере это Тир–тРНКТир с антикодоном АУА, который комплементарен кодону УАУ).
На рибосоме рядом оказываются две аминокислоты, между которыми образуется пептидная связь. Связь между предыдущей аминокислотой и её тРНК (в нашем примере между глицином и тРНКГли) разрывается.
Затем рибосома смещается еще на один триплет, и в результате транслокации тРНК, которая была на Р–участке (в нашем примере тРНКГли), оказывается за пределами рибосомы и отщепляется от мРНК. А–участок освобождается, и рабочий цикл рибосомы начинается сначала.
Терминация. Заключается в окончании синтеза полипептидной цепи.
В конце концов, рибосома достигает такого кодона мРНК, которому не соответствует ни одна тРНК (и ни одна аминокислота). Существует три таких нонсенс–кодона: УАА («охра»), УАГ («янтарь»), УГА («опал»). На этих кодонах мРНК рабочий цикл рибосомы прерывается, и наращивание полипептида прекращается. Рибосома под воздействием определенных белков вновь разделяется на субъединицы.
Модификация белков. Как правило, синтезированный полипептид подвергается дальнейшим химическим превращениям. Исходная молекула может разрезаться на отдельные фрагменты; затем одни фрагменты сшиваются, другие гидролизуются до аминокислот. Простые белки могут соединяться с самыми разнообразными веществами, образуя гликопротеины, липопротеины, металлопротеины, хромопротеины и другие сложные белки. Кроме того, аминокислоты уже в составе полипептида могут подвергаться химическим превращениям. Например, аминокислота пролин, входящая в состав белка проколлагена, окисляется до гидроксипролина. В результате из проколлагена образуется коллаген – основной белковый компонент соединительной ткани.
Реакции модификации белков не являются реакциями матричного типа. Такие биохимические реакции называются ступенчатыми.
Энергетика биосинтеза белков. Биосинтез белков – очень энергоемкий процесс. При аминоацилировании тРНК затрачивается энергия одной связи молекулы АТФ, при кодонзависимом связывании аминоацил-тРНК – энергия одной связи молекулы ГТФ, при перемещении рибосомы на один триплет – энергия одной связи еще одной молекулы ГТФ. В итоге на присоединение аминокислоты к полипептидной цепи затрачивается около 90 кДж/моль. При гидролизе же пептидной связи высвобождается лишь 2 кДж/моль. Таким образом, при биосинтезе большая часть энергии безвозвратно теряется (рассеивается в виде тепла).
Генетический код, его основные свойства
В ходе реакций матричного синтеза на основании генетического кода синтезируется полипептид с наследственно обусловленной структурой. Отрезок ДНК, содержащий информацию о структуре определенного полипептида, называется ген.
Однако, ген – это не просто участок ДНК, а единица наследственной информации, носителем которой являются нуклеиновые кислоты. Установлено, что ген имеет сложную структуру.
В большинстве случаев кодирующие участки (экзоны) разделены некодирующими (интронами). В то же время, благодаря альтернативному сплайсингу, деление участка ДНК на кодирующие и некодирующие оказывается условным. Некоторые участки ДНК могут перемещаться относительно друг друга – их называют мобильными генетическими элементами (МГЭ). Многие гены представлены несколькими копиями – тогда один и тот же белок кодируется разными участками ДНК. Еще сложнее закодирована генетическая информация у вирусов. У многих из них обнаружены перекрывающиеся гены: один и тот же участок ДНК может транскрибироваться с разных стартовых точек.
Процесс экспрессии генов обладает гибкостью: одному участку ДНК может соответствовать несколько полипептидов; один полипептид может кодироваться разными участками ДНК. Окончательная модификация белков происходит с помощью ферментов, которые кодируются различными участками ДНК.
Общие свойства генетического кода
Отражение одних объектов с помощью других называется кодированием. Отражение структуры белков в виде триплетов ДНК называется кодом ДНК, или генетическим кодом. Благодаря генетическому коду устанавливается однозначное соответствие между нуклеотидными последовательностями нуклеиновых кислот и аминокислотами, входящими в состав белков. Генетический код обладает следующими основными свойствами:
1. Генетический код триплетен: каждая аминокислота кодируется триплетом нуклеотидов ДНК и соответствующим триплетом иРНК. При этом кодоны ничем не отделены друг от друга (отсутствуют «запятые»).
2. Генетический код является избыточным (вырожденным): почти все аминокислоты могут кодироваться разными кодонами. Только двум аминокислотам соответствует по одному кодону: метионину (АУГ) и триптофану (УГГ). Зато лейцину, серину и аргинину соответствует по 6 разных кодонов.
3. Генетический код является неперекрывающимся: каждая пара нуклеотидов принадлежит только одному кодону (исключения обнаружены у вирусов).
4. Генетический код един для подавляющего большинства биологических систем. Однако имеются и исключения, например, у инфузорий и в митохондриях разных организмов. Поэтому генетический код называют квазиуниверсальным.
Биосинтез белка и генетический код
Определение 1
Биосинтез белка – ферментативный процесс синтеза белков в клетке. В нём участвуют три структурные элемента клетки – ядро, цитоплазма, рибосомы.
В ядре клетки в молекулах ДНК сохраняется информация о всех белках, которые в ней синтезируются, зашифрованная с помощью четырёхбуквенного кода.
Определение 2
Генетический код – это последовательность расположения нуклеотидов в молекуле ДНК, которая определяет последовательность аминокислот в молекуле белка.
Свойства генетического кода таковы:
Генетический код триплетный, то есть каждой аминокислоте соответствует свой кодовый триплет (кодон ), состоящий из трёх расположенных рядом нуклеотидов.
Пример 1
Аминокислота цистеин кодируется триплетом А-Ц-А, валин – триплетом Ц-А-А.
Код не перекрывается, то есть нуклеотид не может входить в состав двух соседних триплетов.
Код вырожден, то есть одна аминокислота может кодироваться несколькими триплетами.
Пример 2
Аминокислота тирозин кодируется двумя триплетами.
Код не имеет запятых (разделительных знаков), считывание информации происходит тройками нуклеотидов.
Определение 3
Ген – участок молекулы ДНК, который характеризуется определённой последовательностью нуклеотидов и определяет синтез одногой полипептидной цепи.
Код является универсальным, то есть единым для всех живых организмов – от бактерий до человека. У всех организмов есть одни и те же 20 аминокислот, которые кодируются одними и теми же триплетами.
Этапы биосинтеза белка: транскрипция и трансляция
Структура любой белковой молекулы закодирована в ДНК, которая не участвует непосредственно в её синтезе. Она служит лишь матрицей для синтеза РНК.
Процесс биосинтеза белка происходит на рибосомах, которые расположены преимущественно в цитоплазме. Значит, для осуществления передачи к месту синтеза белка генетической информации из ДНК нужен посредник. Эту функцию выполняет иРНК.
Определение 4
Процесс синтеза молекулы иРНК на одной цепи молекулы ДНК на основании принципа комплементарности называется транскрипцией , или переписыванием.
Транскрипция происходит в ядре клетки.
Процесс транскрипции осуществляется одновременно не на всей молекуле ДНК, а лишь на её небольшом участке, который отвечает определённому гену. При этом происходит раскручивание части двойной спирали ДНК и короткий участок одной из цепей оголяется – теперь он будет выполнять роль матрицы для синтеза иРНК.
Потом вдоль этой цепи двигается фермент РНК-полимераза, соединяющий нуклеотиды в цепь иРНК, которая удлиняется.
Замечание 2
Транскрипция может одновременно происходить и на нескольких генах одной хромосомы и на генах разных хромосомах.
Образованная в результате иРНК содержит последовательность нуклеотидов, которая является точной копией последовательности нуклеотидов на матрице.
Замечание 3
Если в молекуле ДНК есть азотистое основание цитозин, то в иРНК – гуанин и наоборот. Комплементарной парой в ДНК является аденин – тимин, а РНК вместо тимина содержит урацил.
На специальных генах синтезируются и два другие типа РНК – тРНК и рРНК.
Начало и окончание синтеза всех типов РНК на матрице ДНК строго фиксированы специальными триплетами, которые контролируют запуск (инициирующие) и остановку (терминальные) синтеза. Они выполняют функции «разделительных знаков» между генами.
Соединение тРНК с аминокислотами происходит в цитоплазме. Молекула тРНК формой напоминает листик клевера, на его верхушке расположен антикодон – триплет нуклеотидов, который кодирует аминокислоту, которую переносит данная тРНК.
Сколько видов аминокислот, столько существует и тРНК.
Замечание 4
Поскольку много аминокислот могут кодироваться несколькими триплетами, то количество тРНК больше 20 (известно около 60 тРНК).
Соединение тРНК с аминокислотами происходит с участием ферментов. Молекулы тРНК транспортируют аминокислоты к рибосомам.
Определение 5
Трансляция – это процесс, в результате которого информация о структуре белка, записанная в иРНК в виде последовательности нуклеотидов, реализуется в виде последовательности аминокислот в молекуле белка, которая синтезируется.
Этот процесс осуществляется в рибосомах.
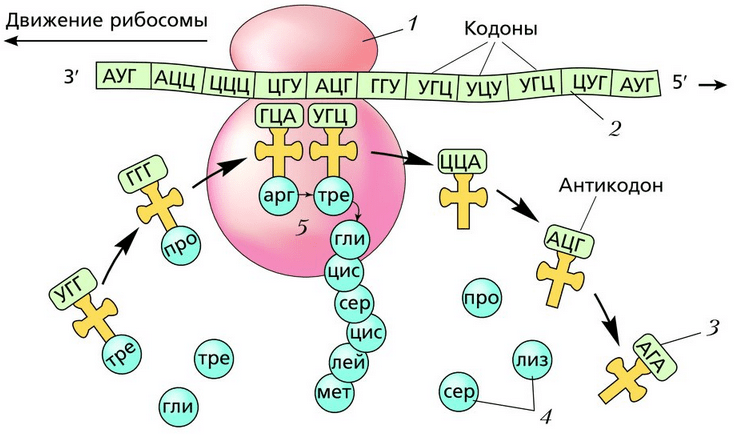
Сначала иРНК присоединяется к рибосоме. На иРНК «нанизывается» первая рибосома, которая синтезирует белок. По мере продвижения рибосомы на конец иРНК, который освободился, «нанизывается» новая рибосома. На одной иРНК могут находиться одновременно более 80 рибосом, которые синтезируют один и тот же белок. Такая группа рибосом, соединённых с одной иРНК, называется полирибосомой , или полисомой . Вид белка, который синтезируется, определяется не рибосомой, а информацией, записанной на иРНК. Одна и та же рибосома способна синтезировать разные белки. После завершения синтеза белка рибосома отделяется от иРНК, а белок поступает в эндоплазматическую сеть.
Каждая рибосома состоит из двух субъединиц – малой и большой. Молекула иРНК присоединяется к малой субъединице. В месте контакта рибосомы и иРН находятся 6 нуклеотидов (2 триплета). К одному из них всё время подходят из цитоплазмы тРНК с разными аминокислотами и касаются антикодоном кодона иРНК. Если триплеты кодона и антикодона оказываются комплементарными, между аминокислотой уже синтезированной части белка и аминокислотой, которая доставляется тРНК, возникает пептидная связь. Соединение аминокислот в молекулу белка осуществляется с участием фермента синтетазы. Молекула тРНК отдаёт аминокислоту и переходит в цитоплазму, а рибосома передвигается на один триплет нуклеотидов. Так последовательно синтезируется полипептидная цепь. Продолжается всё это до тех пор, пока рибосома не дойдёт к одному из трёх терминирующих кодонов: УАА, УАГ или УГА. После этого синтез белка прекращается.
Замечание 5
Таким образом, последовательность кодонов иРНК определяет последовательность включения аминокислот в цепь белка. Синтезированные белки поступают в каналы эндоплазматического ретикулюма. Одна молекула белка в клетке синтезируется за 1 - 2 минуты.
Биосинтез белков является важнейшим процессом анаболизма. Все признаки, свойства и функции клеток и организмов определяются в конечном итоге белками. Белки недолговечны, время их существования ограничено. В каждой клетке постоянно синтезируются тысячи различных белковых молекул. В начале 50-х гг. ХХ в. Ф. Крик сформулировал центральную догму молекулярной биологии: ДНК → РНК → белок. Согласно этой догме способность клетки синтезировать определенные белки закреплена наследственно, информация о последовательности аминокислот в белковой молекуле закодирована в виде последовательности нуклеотидов ДНК. Участок ДНК, несущий информацию о первичной структуре конкретного белка, называетсягеном . Гены не только хранят информацию о последовательности аминокислот в полипептидной цепочке, но и кодируют некоторые виды РНК: рРНК, входящие в состав рибосом, и тРНК, отвечающие за транспорт аминокислот. В процессе биосинтеза белка выделяют два основных этапа: транскрипция - синтез РНК на матрице ДНК (гена) - и трансляция - синтез полипептидной цепи.
Генетический код и его свойства
Генетический код - система записи информации о последовательности аминокислот в полипептиде последовательностью нуклеотидов ДНК или РНК. В настоящее время эта система записи считается расшифрованной.
Свойства генетического кода:
- триплетность: каждая аминокислота кодируется сочетанием из трех нуклеотидов (триплетом, кодоном);
- однозначность (специфичность): триплет соответствует только одной аминокислоте;
- вырожденность (избыточность): аминокислоты могут кодироваться несколькими (до шести) кодонами;
- универсальность: система кодирования аминокислот одинакова у всех организмов Земли;
- неперекрываемость: последовательность нуклеотидов имеет рамку считывания по 3 нуклеотида, один и тот же нуклеотид не может быть в составе двух триплетов;
- из 64 кодовых триплетов 61 - кодирующие, кодируют аминокислоты, а 3 - бессмысленные (в РНК - УАА, УГА, УАГ), не кодируют аминокислоты. Они называются кодонами-терминаторами , поскольку блокируют синтез полипептида во время трансляции. Кроме того, есть кодон-инициатор (в РНК - АУГ), с которого трансляция начинается.
| Первое основание | Второе основание | Третье основание |
|||
|---|---|---|---|---|---|
| У(А) | Ц(Г) | А(Т) | Г(Ц) | ||
| У(А) | Фен Фен Лей Лей | Сер Сер Сер Сер | Тир Тир - - | Цис Цис - Три | У(А) Ц(Г) А(Т) Г(Ц) |
| Ц(Г) | Лей Лей Лей Лей | Про Про Про Про | Гис Гис Глн Глн | Арг Арг Арг Арг | У(А) Ц(Г) А(Т) Г(Ц) |
| А(Т) | Иле Иле Иле Мет | Тре Тре Тре Тре | Асн Асн Лиз Лиз | Сер Сер Арг Арг | У(А) Ц(Г) А(Т) Г(Ц) |
| Г(Ц) | Вал Вал Вал Вал | Ала Ала Ала Ала | Асп Асп Глу Глу | Гли Гли Гли Гли | У(А) Ц(Г) А(Т) Г(Ц) |
* Первый нуклеотид в триплете - один из четырех левого вертикального ряда, второй - один из верхнего горизонтального ряда, третий - из правого вертикального.
Реакции матричного синтеза
Это особая категория химических реакций, происходящих в клетках живых организмов. Во время этих реакций происходит синтез полимерных молекул по плану, заложенному в структуре других полимерных молекул-матриц. На одной матрице может быть синтезировано неограниченное количество молекул-копий. К этой категории реакций относятся репликация, транскрипция, трансляция и обратная транскрипция.
Ген - участок молекулы ДНК, кодирующий первичную последовательность аминокислот в полипептиде или последовательность нуклеотидов в молекулах транспортных и рибосомных РНК. ДНК одной хромосомы может содержать несколько тысяч генов, которые располагаются в линейном порядке. Место гена в определенном участке хромосомы называется локусом . Особенностями строения гена эукариот являются: 1) наличие достаточно большого количества регуляторных блоков, 2) мозаичность (чередование кодирующих участков с некодирующими).
Экзоны (Э) - участки гена, несущие информацию о строении полипептида.
Интроны (И) - участки гена, не несущие информацию о строении полипептида. Число экзонов и интронов различных генов разное; экзоны чередуются с интронами, общая длина последних может превышать длину экзонов в два и более раз. Перед первым экзоном и после последнего экзона находятся нуклеотидные последовательности, называемые соответственно лидерной (ЛП) и трейлерной последовательностью (ТП). Лидерная и трейлерная последовательности, экзоны и интроны образуют единицу транскрипции.
Промотор (П) - участок гена, к которому присоединяется фермент РНК-полимераза, представляет собой особое сочетание нуклеотидов. Перед единицей транскрипции, после нее, иногда в интронах находятся регуляторные элементы (РЭ), к которым относятся энхансеры и сайленсеры . Энхансеры ускоряют транскрипцию, сайленсеры тормозят ее.
Транскрипция - синтез РНК на матрице ДНК. Осуществляется ферментом РНК-полимеразой.
РНК-полимераза может присоединиться только к промотору, который находится на 3′-конце матричной цепи ДНК, и двигаться только от 3′- к 5′-концу этой матричной цепи ДНК. Синтез РНК происходит на одной из двух цепочек ДНК в соответствии с принципами комплементарности и антипараллельности. Строительным материалом и источником энергии для транскрипции являются рибонуклеозидтрифосфаты (АТФ, УТФ, ГТФ, ЦТФ).
В результате транскрипции образуется «незрелая» иРНК (про-иРНК), которая проходит стадию созревания или процессинга. Процессинг включает в себя: 1) КЭПирование 5′-конца, 2) полиаденилирование 3′-конца (присоединение нескольких десятков адениловых нуклеотидов), 3) сплайсинг (вырезание интронов и сшивание экзонов). В зрелой иРНК выделяют КЭП, транслируемую область (сшитые в одно целое экзоны), нетранслируемые области (НТО) и полиадениловый «хвост».
Транслируемая область начинается кодоном-инициатором, заканчивается кодонами-терминаторами. НТО содержат информацию, определяющую поведение РНК в клетке: срок «жизни», активность, локализацию.
Транскрипция и процессинг происходят в клеточном ядре. Зрелая иРНК приобретает определенную пространственную конформацию, окружается белками и в таком виде через ядерные поры транспортируется к рибосомам; иРНК эукариот, как правило, моноцистронны (кодируют только одну полипептидную цепь).
Трансляция
Трансляция - синтез полипептидной цепи на матрице иРНК.
Органоиды, обеспечивающие трансляцию, - рибосомы. У эукариот рибосомы находятся в некоторых органоидах - митохондриях и пластидах (70S-рибосомы), в свободном виде в цитоплазме (80S-рибосомы) и на мембранах эндоплазматической сети (80S-рибосомы). Таким образом, синтез белковых молекул может происходить в цитоплазме, на шероховатой эндоплазматической сети, в митохондриях и пластидах. В цитоплазме синтезируются белки для собственных нужд клетки; белки, синтезируемые на ЭПС, транспортируются по ее каналам в комплекс Гольджи и выводятся из клетки. В рибосоме выделяют малую и большую субъединицы. Малая субъединица рибосомы отвечает за генетические, декодирующие функции; большая - за биохимические, ферментативные.
В малой субъединице рибосомы расположен функциональный центр (ФЦР) с двумя участками - пептидильным (Р-участок) иаминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три - в пептидильном и три - в аминоацильном участках.
Для транспорта аминокислот к рибосомам используются транспортные РНК, тРНК. Длина тРНК от 75 до 95 нуклеотидных остатков. Они имеют третичную структуру, по форме напоминающую лист клевера. В тРНК различают антикодоновую петлю и акцепторный участок. В антикодоновой петле РНК имеется антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3′-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединять именно эту аминокислоту (с затратой АТФ). Таким образом, у каждой аминокислоты есть свои тРНК и свои ферменты, присоединяющие аминокислоту к тРНК.
Двадцать видов аминокислот кодируются 61 кодоном, теоретически может быть 61 вид тРНК с соответствующими антикодонами. Но кодируемых аминокислот всего 20 видов, значит, у одной аминокислоты может быть несколько тРНК. Установлено существование нескольких тРНК, способных связываться с одним и тем же кодоном (последний нуклеотид в антикодоне тРНК не всегда важен), поэтому в клетке обнаружено всего около 40 различных тРНК.
Синтез белка начинается с того момента, когда к 5′-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК (транспортирующая аминокислоту метионин). Следует отметить, что любая полипептидная цепь на N-конце сначала имеет метионин, который в дальнейшем чаще всего отщепляется. Синтез полипептида идет от N-конца к С-концу, то есть пептидная связь образуется между карбоксильной группой первой и аминогруппой второй аминокислот.
Затем происходит присоединение большой субъединицы рибосомы, и в А-участок поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке.
Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ.
Как только образовалась пептидная связь, метиониновая тРНК отсоединяется от метионина, а рибосома передвигается на следующий кодовый триплет иРНК, который оказывается в А-участке рибосомы, а метиониновая тРНК выталкивается в цитоплазму. На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами.
Трансляция идет до тех пор, пока в А-участок не попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения. Полипептидная цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субъединиц рибосомы.
Скорость передвижения рибосомы по иРНК - 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут. Первым белком, синтезированным искусственно, был инсулин, состоящий из 51 аминокислотного остатка. Потребовалось провести 5000 операций, в работе в течение трех лет принимали участие 10 человек.
В трансляции можно выделить три стадии: а) инициации (образование иницаторного комплекса), б) элонгации (непосредственно «конвейер», соединение аминокислот друг с другом), в) терминации (образование терминирующего комплекса).
Транскрипция и трансляция у прокариот
«Механизмы» сборки полинуклеотидных и полипептидных цепочек у прокариот и эукариот не различаются. Но в связи с тем, что гены прокариот не имеют экзонов и интронов (исключение - гены архебактерий), располагаются группами, и на эту группу генов приходится один промотор, появляются следующие особенности транскрипции и трансляции у прокариот.
- В результате транскрипции образуется полицистронная иРНК, кодирующая несколько белков, совместно обеспечивающих определенную группу реакций.
- иРНК имеет несколько центров инициации трансляции, терминации трансляции и НТО.
- Не происходят КЭПирование, полиаденилирование и сплайсинг иРНК.
- Трансляция начинается еще до завершения транскрипции; эти процессы не разделены во времени и пространстве, как это имеет место у эукариот.
Можно добавить, что срок «жизни» прокариотических иРНК - несколько минут (у эукариот - часы и даже сутки).
Если видео не работает. Дополнительный
Наиболее важным процессом пластического обмена является биосинтез белка. Он протекает во всех клетках организмов.
Генетический код. Аминокислотная последовательность в молекуле белка зашифрована в виде нуклеотидной последовательности в молекуле ДНК и называется генетическим кодом. Участок молекулы ДНК, ответственный за синтез одного белка, называется геном.
Характеристика генетического кода.
1. Код триплетен: каждой аминокислоте соответствует сочетание из 3 нуклеотидов. Всего таких сочетаний - 64 кода. Из них 61 код смысловой, т. е. соответствует 20 аминокислотам, а 3 кода - бессмысленные, стоп-коды, которые не соответствуют аминокислотам, а заполняют промежутки между генами.
2. Код однозначен - каждый триплет соответствует только одной аминокислоте.
3. Код вырожден - каждая аминокислота имеет более чем один код. Например, у аминокислоты глицин - 4 кода: ЦЦА, ЦЦГ, ЦЦТ, ЦЦЦ, чаще у аминокислот их 2-3.
4. Код универсален - все живые организмы имеют один и тот же генетический код аминокислот.
5. Код непрерывен - между кодами нет промежутков.
6. Код неперекрываем - конечный нуклеотид одного кода не может служить началом другого.
Условия биосинтеза
Для биосинтеза белка необходима генетическая информация молекулы ДНК; информационная РНК - переносчик этой информации из ядра к месту синтеза; рибосомы - органоиды, где происходит собственно синтез белка; набор аминокислот в цитоплазме; транспортные РНК, кодирующие аминокислоты и переносящие их к месту синтеза на рибосомы; АТФ - вещество, обеспечивающее энергией процесс кодирования и биосинтеза.
Этапы
Транскрипция - процесс биосинтеза всех видов РНК на матрице ДНК, который протекает в ядре.
Определенный участок молекулы ДНК деспирализуется, водородные связи между двумя цепочками разрушаются под действием ферментов. На одной цепи ДНК, как на матрице, по принципу комплементарное из нуклеотидов синтезируется РНК-копия. В зависимости от участка ДНК таким образом синтезируются рибосомные, транспортные, информационные РНК.
После синтеза иРНК она выходит из ядра и направляется в цитоплазму к месту синтеза белка на рибосомы.
Трансляция - процесс синтеза полипептидных цепей, осуществляемый на рибосомах, где иРНК является посредником в передаче информации о первичной структуре белка.
Биосинтез белка состоит из ряда реакций.
1. Активирование и кодирование аминокислот. тРНК имеет вид клеверного листа, в центральной петле которого располагается триплет-ный антикодон, соответствующий коду определенной аминокислоты и кодону на иРНК. Каждая аминокислота соединяется с соответствующей тРНК за счет энергии АТФ. Образуется комплекс тРНК-аминокислота, который поступает на рибосомы.
2. Образование комплекса иРНК-рибосома. иРНК в цитоплазме соединяется рибосомами на гранулярной ЭПС.
3. Сборка полипептидной цепи. тРНК с аминокислотами по принципу комплементарности антикодона с кодоном соединяются с иРНК и входят в рибосому. В пептидном центре рибосомы между двумя аминокислотами образуется пептидная связь, а освободившаяся тРНК покидает рибосому. При этом иРНК каждый раз продвигается на один триплет, внося новую тРНК - аминокислоту и вынося из рибосомы освободившуюся тРНК. Весь процесс обеспечивается энергией АТФ. Одна иРНК может соединяться с несколькими рибосомами, образуя полисому, где идет одновременно синтез многих молекул одного белка. Синтез заканчивается, когда на иРНК начинаются бессмысленные кодоны (стоп-коды). Рибосомы отделяются от иРНК, с них снимаются полипептидные цепи. Так как весь процесс синтеза протекает на гранулярной эндо-плазматической сети, то образовавшиеся полипептидные цепи поступают в канальца ЭПС, где приобретают окончательную структуру и превращаются в молекулы белка.
Все реакции синтеза катализируются специальными ферментами с затратой энергии АТФ. Скорость синтеза очень велика и зависит от длины полипептида. Например, в рибосоме кишечной палочки белок из 300 аминокислот синтезируется приблизительно за 15-20 с.
Биосинтез белка - сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков, происходящий на рибосомах клеток живых организмов с участием молекул мРНК и тРНК.
ББ протекает в два этапа – транскрипция (от ДНК до синтеза зрелой мРНК), трансляция (с выхода зрелой мРНК в цитоплазму и синтеза полипептида.)
Трансляция. В прокариотических клетках процесс трансляции сопряжен с синтезом мРНК: они происходят практически одновременно.
Белки в рибосоме держатся на каркасе, состоящем из рибосомной РНК. Формирование рибосомы начинается с того, что рибосомная РНК сворачивается и на нее в определенном порядке начинают налипать белки. На рисунке представлена рибосомная РНК. В ней самокомплементарные участки нити РНК спариваются, образуя шпильки (вторичная структура), и затем РНК сворачивается (третичная структура РНК), образуя каркас субчастиц.
Еще один вид РНК, участвующей в синтезе белка, это транспортная РНК (тРНК). Молекулы тРНК относительно небольшие (по сравнению с рибосомной или матричной РНК). Все тРНК имеют общую вторичную структуру. За счет спаривания комплементарных участков молекулы тРНК образуется три "стебля" с петлями на концах и один "стебель", образованный 5"- и 3"-концами молекулы тРНК (иногда образуется еще дополнительная пятая петля). Изображение этой структуры похоже на крест или клеверный лист. "Голова" на этом листе представлена антикодонной петлей, здесь находится антикодо – те три нуклеотида, которые комплементарно взаимодействуют с кодоном в мРНК. Противоположный антикодонной петле стебель, образованный концами молекулы, называется акцепторным стеблем – сюда присоединяется соответствующая аминокислота. Распознают подходящие друг другу тРНК и аминокислоты специальные ферменты, называемые аминоацил-тРНК синтетазами. Для каждой аминокислоты есть своя аминоацил-тРНК синтетаза.
В рибосоме находится матричная РНК (мРНК). С кодоном (тремя нуклеотидами) мРНК комплементарно связан антикодон транспортной РНК, на которой висит остаток аминокислоты. На рисунке видна такая структура (тРНК вместе с аминокислотой, которая называется аминоцил-тРНК).
Процесс трансляции, также как и процесс транскрипции, связан с перемещением вдоль молекулы нуклеиновой кислоты, разница в том, что рибосома шагает на три нуклеотида, в то время как РНК-полимераза - на один.
Рибосомы про- и эукариот очень сходны по структуре и функциям. Они состоят из двух субчастиц: большой и малой. У эукариот малая субчастица образована одной молекулой рРНК и 33 молекулами разных белков. Большая субчастица объединяет три молекулы рРНК и около 40 белков. Прокариотические рибосомы и рибосомы митохондрий и пластид содержат меньше компонентов.
15. Этапы трансляции (инициации, элонгации, терминации)
Инициация трансляции . Синтез белка в большинстве случаев начинается с AUG-кодона, кодирующего метионин. Этот кодон обычно называют стартовым или инициаторным. Инициация трансляции предусматривает узнавание рибосомой этого кодона и привлечение инициаторной аминоацил-тРНК. Для инициации необходимо также наличие определённых нуклеотидных последовательностей в районе стартового кодона. Немаловажн. роль в защите 5"-конца мРНК принадлеж. 5"-кэпу. Сущ. последовательности, отличающей стартовый AUG от внутренних совершенно необходимо, так как в противном случае инициация синтеза белка происходила бы хаотично на всех AUG-кодонах. Процесс инициации обеспеч. специальными белками - факторами инициации. (кот. подвижно связаны с малой субчастицей рибосомы. По завершении фазы инициации и образования комплекса рибосома - мРНК - инициирующая аминоацил-тРНК эти факторы отделяются от рибосомы.) Механизмы инициации трансляции у про- и эукариот существенно отличаются: прокариотические рибосомы потенциально способны нах. стартовый AUG и инициировать синтез на любых участках мРНК.
Фаза элонгации , или удлинения пептида, включает в себя все реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. Она представляет собой циклически повторяющиеся события, при которых происходит специфическое узнавание аминоацил-тРНК очередного кодона, находящегося в А-участке, комплементарное взаимодействие между антикодоном и кодоном. В процессе наращивания полипептидной цепи принимают участие два белковых фактора элонгации. Первый (EF1a у эукариот, EF-Tu - у прокариот) переносит заряженную тРНК в А (аминоацил)-сайт рибосомы. После формирования пептидной связи, что катализируется рРНК, и переноса связанной с тРНК пептида в из Р-сайта в А-сайт второй белок (EF2 у эукариот, EF-G - у прокариот) катализирует перемещение рибосомы на один триплет. Таким образом петидил-тРНК оказывается вновь в Р-сайте, а «пустая» тРНК в Р-сайте - в Е-сайте. Цикл элонгации завершается, когда новая тРНК с антикодоном, подходящим к кодону в А-сайте доставлена EF1a (или EF-Tu). Сборка пептидной цепи осуществляется с достаточно большой скоростью, зависящей от температуры. У бактерий при 37 °С она выражается в добавлении к подипептиду от 12 до 17 аминокислот в 1 с. В эукариотических клетках эта скорость ниже и выражается в добавлении двух АК в 1 с.
Фаза терминации, или завершения синтеза полипептида, связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ или У ГА), когда тот входит в зону А-участка рибосомы. При этом к последней аминокислоте в пептидной цепи присоединяется вода, и ее карбоксильный конец отделяется от тРНК. В результате завершенная пептидная цепь теряет связь с рибосомой, кот. распадается на две субчастицы.